Theory and tradition suggest that when individuals are raised in environments where kinship is emphasized, the likelihood of sexual relationships diminishes, thereby enhancing safety for children. Conversely, when children are cared for by strangers—where these protective relationship-mechanisms are absent—there may be increased vulnerability to sexual exploitation.
Video and text of Sheila Newman's speech at the Animal Justice Party's event, "Policy basis for Kangaroo treatment in the ACT," 5 April 2016: Harvesting, damage mitigation and culling probably actually accelerate population growth in roos because the smaller ones survive and adapt by sexually maturing earlier - which speeds up fertility turnover. Since 2003 DNA studies have shown that ACT and southern NSW roos, both male and female, migrate at significant rates and for longer distances than the ACT model assumes. Migration has probably been mistaken for fertility, rendering ACT roo counts unreliable and invalid. The ACT needs to stop culling and widen its research base to consider various genetically based algorithms that naturally restrain fertility opportunities in kangaroos.
Examples include separate gender pathways, with 'sexual segregation' where male and female populations live apart. It is likely that the stable presence of mature dominant males and females in family and mob organisation inhibits sexual maturity and activity as has been shown in studies of other species, such as macaques and superb fairy wrens (the latter cooperative breeders). In humans, girls brought up with step-fathers who came late to the family were more likely to mature sexually earlier due to absence of Westermarck Effect.)
Planned wildlife corridors need to be made safe and long-term viable to cope with people, car and kangaroo population movements.
Canberra is pursuing a policy of rapid population growth, mostly through invited economic immigration.
Canberra's population problem
In June 2016 ACT - South West Australian Capital Territory was the fastest growing area in Australia and grew by 127.3%. (ABS http://www.abs.gov.au/ausstats/[email protected]/mf/3218.0)
Canberra’s population could increase to 904,000 by 2061 according to new projections released by the Australian Bureau of Statistics. It's not inevitable, but the government would like you to believe it is.
Predicting a population growth of at least 98 per cent within 50 years, ACT population projections for 2061 suggest that the Australian Capital Territory population could exceed Tasmania's population by 2038.
But some think that the ACT's biggest problem is its kangaroo population
It's not the new suburbs, the new roads, the new airport, the additional schools, hospitals, houses, and all the new cars that threaten Canberra's grasslands: it's the eastern grey kangaroos.
It seems that it is better to have cattle in Canberra's nature parks than kangaroos.
Ecological cattle grazing is now being trialed. Cattle can be more easily moved than kangaroos. (Fletcher, Senior Ecologist, communication to P. Machin.)
Although, Fletcher had previously described the devastation cattle made to grass cover in no uncertain terms: "Fletcher Phd: p.37. “70 pregnant cows and four bulls grazed for ten weeks at Tidbinbilla after a bushfire in January 2003 (Section 3.5.1). Prior to their arrival, there had been an atypical abundance of pasture due to the death of almost half of the Tidbinbilla kangaroos in the bushfire, but by the time the cattle were removed, the Tidbinbilla pasture had been reduced to the lowest herbage mass recorded on any site during the study.”
"
ACT Kangaroo Management Policy works on a model that all creatures maximize their population growth and that Canberra's roos are riding an expansive curve which can only be capped by massive frequent culls. A stated fear is that they will otherwise graze and drastically modify biodiversity of Canberra's grasslands. Another is that roos need periodically to be shot so as to save them from starving to death.
Why culling is better than harvesting (ACT Senior Ecologist, Don Fletcher)
"[…] the model indicates that commercial harvesting (currently under trial in the region, at the maximum level allowed, results in a sustainable harvest of kangaroos, but does not increase the herbage mass, and only slightly reduces the frequency of crashes when herbage mass falls to low levels. (To demonstrate this with an ecological experiment would require an extremely large investment of research effort.)
However, an alternative 'national park damage mitigation' formula, which holds kangaroo density to about 1 ha -1 , is predicted to increase herbage mass considerably and to reduce the frequency of crashes in herbage mass, but these effects would be achieved at the cost of having to shoot large numbers of kangaroos." (Fletcher Phd: Population dynamics of Eastern Grey Kangaroos in Temperate Grasslands, 2006, p. vi.)
The model and the reality
The ACT Roo Management model is one of high fertility sedentary populations that rarely migrate, grazing grasslands down to the subsoil. But, in his 2006 thesis, p. 237, Senior ACT Kangaroo management ecologist, Donald Fletcher, tested this model and found, to his surprise, that,
"The study did not provide evidence that high densities of kangaroos reduce groundcover to the levels where erosion can accelerate.
Unmanaged kangaroo populations did not necessarily result in low levels of ground cover. Groundcover had a positive but not significant relationship to kangaroo density, with the highest cover at the wettest site where kangaroo density was highest. Weather has an important influence on groundcover."
"The results from the study as a whole indicate that unmanaged kangaroo populations did not necessarily result in unacceptably low levels of ground cover." (Fletcher Phd: p.231.
The Kangaroo Migration factor
Since 2003 DNA studies have shown that ACT and southern NSW roos, both male and female, migrate at significant rates and for longer distances than the ACT model assumes.
Migration has probably been mistaken for fertility, rendering ACT kangaroo counts unreliable and probably invalid.
DNA studies 2003 show migration a strong factor
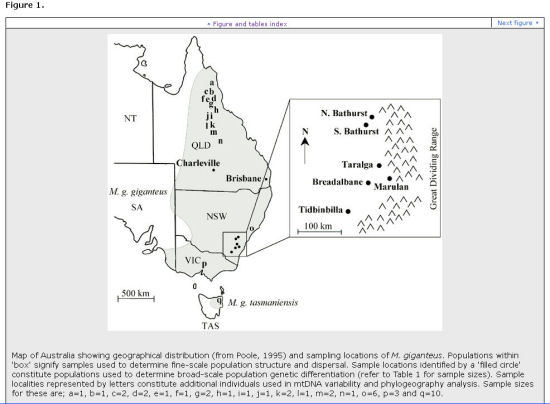
Zenger et al (2003)[1] found that mitrochondrial DNA samples indicated about 22.61 individuals per generation migrated with a range of 8.17-59.30. In female immigrants the range was 2.73 with a range of 0.60-12.16. Although females demonstrate smaller migration rates compared to the sexes combined, the values are still comparatively high. Analysis across NSW showed populations separated by up to about 230km had equivalent numbers of close relatives when compared to populations only about 20km apart.
This contradicted field study opinion that migration was low in eastern grey kangaroos, and especially low in females in the ACT. Tidbinbilla (a Canberra nature park studied by Fletcher) featured in the Zenger et al study.
Zenger MtDNA findings contrary to Migration views in Fletcher thesis
"Throughout their lives eastern grey kangaroos are relatively sedentary (Johnson 1989) compared to red kangaroos (Priddel 1987). A partly concurrent study of eastern grey kangaroo habitat use and movements on the Googong and Tidbinbilla sites found the eastern grey kangaroos on these sites were sedentary in all seasons (minimum convex polygon mean size 0.43 km 2 ± 0.06 SE and 0.61 km 2 ± 0.08 respectively; Viggers and Hearn 2005). Kangaroos were not radio tracked at Gudgenby but my observations suggest there is no more movement of eastern grey kangaroos on and off the site there than at Tidbinbilla. Thus it is likely there was little net movement of kangaroos on and off the study sites." (Fletcher, page v.)
Paradoxical impact of Culls, kills and Harvests
Harvesting, damage mitigation and culling probably actually accelerate population growth in roos because the smaller ones survive and adapt by sexually maturing earlier - which speeds up fertility turnover.
"Smaller, earlier breeding genetic stock tend to escape harvesting". See, e.g. J.J. Poosa, A. Brannstrom, U. Dieckmann, “Harvest-induced maturation evolution under different life-history trade-offs and harvesting regimes.” (See note for more literature on this.)[2]
Fletcher, on estimates of biomass consumption per roo allows for large variations in harvested populations vs wild populations.
"How big are eastern grey kangaroos?
The mean live weight of eastern grey kangaroos taken from the unshot population at Tidbinbilla was 29 kg – smaller than the 35 kg mean live weight assumed in the Kinchega kangaroo study (Caughley et al. 1987). Based on the size relationship between shot and unshot populations of kangaroos in South Australia and Queensland (Grigg 2000), the mean size of eastern grey kangaroos in equivalent shot populations was predicted to be 17 kg live weight. The minimum dressed size accepted by operators of commercial chillers is 17 kg, implying that many of the kangaroos in shot populations (on rural properties) in the ACT region are too small to attract commercial shooters."(Fletcher, p.242.)
Culling has a similar effect.
Earlier maturation would contribute to higher population growth rates. What role does harvesting, culling and farm mitigation killing play in accelerating breeding rates?
“Smaller, eat less, more numerous, more fecund", reproduce earlier
“The management implications arising from this study are numerous and a full account would require a separate report. As one example, kangaroos in these temperate grasslands are on average smaller, eat less, are more numerous, and are more fecund, than would be predicted
from other studies (e.g. Caughley et al. 1987). Thus the benefit of shooting each kangaroo, in terms of grass production, is less, or, in other words, more kangaroos have to be shot to achieve a certain level of impact reduction, and the population will recover more quickly, than would have been predicted prior to this study.” (Fletcher, p245.)
Kangaroos shot in Tidbinbilla and low weight in shot populations
“The mean live weight of eastern grey kangaroos in high density populations can be estimated from the weights of a sample of 332 kangaroos shot at Tidbinbilla in June 1997 (Graeme Coulson, personal communication, 2003) to be 29 kg. (That is an adjustment of the actual mean liveweight of the shot sample, 26.4 kg, to allow for seasonal effects, as explained in Discussion. Kangaroos in shot populations, such as on grazing properties, are likely to be smaller due to selective harvesting, also explained in Discussion).”(Fletcher p.242.)
Wider Research Base needed
In my view, the ACT needs to stop culling and widen its research base to consider encouraging various genetically based behaviours that naturally restrain fertility opportunities in roos.
Known examples include
incest avoidance, which limits breeding unless animals can disperse to their own territory. [3] Sexual segregation and gender pathways, where male and female populations live apart.
Incest avoidance as a spatial limiter of breeding opportunity
Many examples of suppressed maturity or breeding in both males and females close by related adults in many species. (Sheila Newman, Demography Territory Law: Rules of animal and human populations, Countershock Press, 2013, chapter 3.) (Paperback edition and Kindle edition.)
In kangaroos male sexual dominance and monopolisation of females is a very obvious trait. (The effect of dominant close females on female maturation is less known and should be investigated as it has been in other species).
Where large males and females are removed from mobs, these limiting population effects are also removed. What happens then?
Kinship rules and incest avoidance
The following diagram is of human kinship rules, however similar patterns of incest avoidance occur in other species, and in kangaroos. The diagram for humans is split into family and in-laws and sets out some typical rules for incest avoidance in low fertility environments - central Australia and mountainous South Korea. The rules for inlaws are reproduced back to front to demonstrate a mirror-like effect. The person in the top left corner, 'You" may not conceive/marry any of the people in the black squares. That leaves only eight possible mates - as long as they are not already married. Imagine how hard it would be to find a wife or husband under these circumstances in a sparsely populated society of small clans that only travelled on foot, without cars, planes or boats. In a more fertile environment, the rules of incest avoidance are usually much less strict, as in Leviticus, where people may marry their first cousins - giving much greater fertility opportunities, even without the benefit of modern transport. For more about this and how it affects human economies see: "Overpopulation: Endogamy,Exogamy and fertility opportunity theory"
The following two diagrams are from Zenger et al (2003).
They show the regions from which their eastern grey kangaroo DNA samples were taken, and they give a 'family tree' of roo DNA diversity, which shows greatly decreased diversity in north NSW and in Queensland. The authors could find no explanation for this.
Possible explanation for decreased Mt DNA in Queensland & Northern NSW
Harvesting has gone on for a long time in these regions. We know it is associated with marked size decrease. It seems likely that it is also associated with earlier sexual maturity. Consider the possibility that, as well as size decrease and earlier sexual maturation in harvested populations, the decrease in genetic diversity present in those populations may be due to inbreeding resulting from loss of family structure and associated incest avoidance, with decreased migration as small early maturing roos settle for their siblings. There seems little will to investigate this. Although there is some literature, it is very limited. (See note [2].)
Sexual Segregation/Gender pathways
Review of Scientific Literature Relevant to the Commercial Harvest Management of Kangaroos http://www.environment.nsw.gov.au/resources/nature/110641Kangaroolitreview.pdf
“Sexual segregation is a phenomenon seen in many species, with segregation occurring along behavioural or ecological dimensions. Sexual segregation in western grey and red kangaroos in semi - arid Victoria has been the subject of intensive investigations since the last review.
[…] MacFarlane and Coulson (2005) investigated the effects of mating activity, group […] composition, spatial distribution and habitat selection on sexual segregation in western grey and red kangaroos. The synchrony and timing of mating activity was seen to influence the magnitude and timing of social segregation in these species, with mixed sex groups predominating during the breeding season. …
… Spatial segregation and habitat segregation were also seen. Although the magnitude of these types of segregation were weaker, they were both still significantly influenced by synchrony and timing of breeding.
Coulson et al. (2006) discussed sexual segregation at three levels (habitat, social and dietary) and confirmed that both size and sex influence segregation.
MacFarlane and Coulson (2009) showed that the need for males to maintain contact with other males (perhaps to develop important fighting skills, evaluate rivals and establish a dominance hierarchy) might also promote sexual segregation.
Similarly Nave (2002) reported evidence of sexual segregation in eastern grey kangaroos in Victoria.”
What are the consequences of loss of sex-specific territory?
Years ago a man who had worked in PNG told me that fertility shot up when churches convinced men and women to cohabit, where previously they had separate land and houses.
What effect could reduction of habitat, forced cohabitation, forcibly changed migration routes and wiped out populations have on male/female kangaroo territory and consequently on fertility opportunities?
How do we know that the female bias (recorded by Fletcher) at Tinbinburra, for instance, is not due to that area being female territory?
Female elder kangaroos
Daughters seem to learn from their mothers to look after joeys. Where female kangaroos are early orphaned their parenting skill may increase risks in joey upbringing. The extraordinary rates of joey mortality may have something to do with this. (See Faces in the Mob for a study of success and failure in raising joeys in one mob.)
In conclusion, regarding ACT Kangaroo management:
It seems that ACT Roo Management Policy and Science:
- Fails to monitor family structure (spatial population monitoring)
- Fails to deal with size reduction, fertility increases probably related to culls etc
- Fails to look at behaviour; notably breeding limitations exerted through incest avoidance/dominance and separate male/female territory
- Underestimates immigration (See Zenger et al)
- Fails to use DNA monitoring to help in the above
- Seems excessively presumptive and mechanistic
NOTES
[1] Zenger et al DNA study 2003: (Heredity (2003) 91, 153–162. doi:10.1038/sj.hdy.6800293, K R Zenger, M D B Eldridge and D W Cooper, "Intraspecific variation, sex-biased dispersal and phylogeography of the eastern grey kangaroo (Macropus giganteus)."
http://www.nature.com/hdy/journal/v91/n2/full/6800293a.html)
[2] Harvesting impact literature: Many of these studies arise from fish stocks. Articles quoting studies for kangaroos tend to quote from the same very small amount of literature and to draw equivocal conclusions, frequently paraphrasing each other. Peter T. Hale, "Genetic effects of kangaroo harvesting", Australian Mammalogy 26:75-86 (2004)http://citeseerx.ist.psu.edu/viewdoc/download?doi=10.1.1.319.7936&rep=rep1&type=pdf seems to be the main work cited, but relies on studies which Fletcher's Phd calls into question, has little to say about eastern grey kangaroos but seems to infer that they have similar rates of starvation attributed to red kangaroos.
In Review of Scientific Literature Relevant to the Commercial Harvest Management of Kangaroos (2011) http://www.environment.nsw.gov.au/resources/nature/110641Kangaroolitreview.pdf, pp.27-28, after flagging the potential impact of harvesting on kangaroos, the study concludes with a mere opinion that the impact of size and other harvesting selection on kangaroos probably would not be great, on the assumption that the harvested populations are not isolated. This is pretty much as Hale's study (above) concludes. However we know that the harvested populations in northern NSW and Queensland are genetically isolated and impoverished according to Zenger et all (2003) cited above. Furthermore, the review showed it was aware of Zenger et al.
"The last two reviews concluded that there was no evidence, or potential, that commercial harvesting could alter the genetic structure of kangaroo populations at current harvesting levels (Olsen and
Braysher 2000, Olsen and Low 2006). It was perceived that kangaroo populations would have to be
reduced to very low levels for genetic impacts to become significant (Olsen and Braysher 2000).
Moreover, at the time of the last review, it was concluded that there was an absence of theoretical,
empirical and modelled evidence of genetic impacts at current levels of harvesting” (Olsen and Low
2006, p50) and there were few, if any,examples of harvest‐induced body size selection in terrestrial
vertebrates. While there have not been any studies specifically investigating the potential genetic
impacts of harvesting kangaroos since the last review, there have been a large number of original
research and review papers addressing this question in a range of other vertebrate species,
highlighting the perception that the potential genetic consequences of harvesting may be significant
The human harvest of wild animals is generally not a random process, with harvesters often
selecting phenotypically desirable animals, e.g. those with a large body size or elaborate weaponry,
such as antlers. This therefore has the potential to impose selective pressure on wild populations,
which may result in an alteration to population structure by reducing the frequency of these
desirable phenotypes and/or an overall loss of genetic variation (Allendorf et al. 2008).
Allendorf and Hard (2009) have termed this process “unnatural selection”, which is defined as undesirable changes in an exploited population due to selection against desirable phenotypes. Cited examples of the
effects of selective harvesting on desirable phenotypes include an increase in the number of tuskless elephants (Loxodonta africana) in South Luanga National Park, Zambia, and a decrease in horn size
of bighorn sheep (Ovis canadensis) because of trophy hunting (reviewed in Allendrof and Hard
2009). In the case of bighorn sheep, the observed genotypic and phenotypic effects resulted from
selective harvesting of young males with rapidly growing horns a trait linked with high reproductive
success) before they reached an age where they could achieve high reproductive success (Coltman
et al. 2003). This study highlights the importance of understanding age-specific trait size, rather
than trait size per se.
A recent review by Mysterud (2011) discusses the relative importance of various biotic and abiotic
factors that determine the potential for selective pressure from harvesting. In particular, Mysterud
highlights the importance of assessing selective harvesting within the context of management
regulations, hunting methods, animal trait variance, behaviour and abundance. Mysterud argues that in many cultures large mammal harvesting is not expected to produce strong directional
selection in trait size.
Although many of the factors discussed are of greater relevance to traditional
sport hunting, this review highlights the importance of a number of factors relevant to the
commercial harvesting of kangaroos in Australia.
There is certainly evidence for selective harvesting of larger/older animals within kangaroo
populations, primarily because the economic performance of kangaroo harvesting enterprises is
highly sensitive to variations in average carcase weight (Stayner 2007). Between 1997 and 2009 the
total harvest in NSW comprised between 70 and 89% males. In the case of wallaroos, the
commercial take is even more strongly biased towards males (almost 90%), because females rarely
reach the minimum size dictated by licence and market conditions (Payne 2011). Despite the
preference for larger males, it was reported that harvesters target a range of sizes above the
minimum, especially when densities are reduced and there are fewer target animals (Payne 2011).
There average weight of harvested animals supports this assertion [(Table 2)].
As reported in the last review (Olsen and Low 2006), studies on the potential effects of size-selective harvesting in kangaroos concluded that although there was potential for genetic consequences of
harvesting within a closed population (Tenhumberg et al. 2004), the degree of mobility and
geographic range of genetic populations of kangaroos would be sufficient to ensure that any
localised effects could be countered by immigration (Hale 2004). So, the question remains: does the
recent literature on this topic provide any basis for changing the previous conclusions?
Probably not.
In the big horn sheep example referred to above, the extent of selective harvesting pressure was
probably much stronger than occurs in kangaroo populations. In addition, the population was small,
isolated and had restricted potential for immigration (Coltman et al. 2003), thereby exhibiting
characteristics akin to a closed population. As such, this probably represents a more extreme
example, where prevailing management and biological factors combined to create strong selective
pressure."
Penny Olsen and Tim Low, "Situation Analysis Report, Update on Current State of Scientific Knowledge on Kangaroos in the Environment, Including Ecological and Economic Impact and Effect of Culling," School of Botany and Zoology, Australian National University, Canberra, ACT and 6 Henry Street, Chapel Hill, Queensland, Prepared for the Kangaroo Management Advisory Panel, March 2006
Proceedings of the 2010 RSPCA Australia Scientific Seminar: Convergence or conflict: animal welfare in wildlife management and conservation, Tuesday 23 February 2010, CSIRO Discovery Centre, Canberra https://www.rspca.org.au/files/website/The-facts/Science/Scientific-Seminar/2010/SciSem2010-Proceedings.pdf
[3] Family structure/westermarck/incest avoidance/endogamy/exogamy: Sheila Newman, Demography, Territory, Law: Rules of animal and human populations, Countershock Press, 2013, Chapter 3, “CHAPTER 3: The urge to disperse: Why children don’t usually marry their parents.” (Available amazon.com) Examples of incest avoidance citations within:
"Several studies have shown that maternal relatives avoid mating with one another (rhesus macaques: Smith, 1995; red colobus, Procolobus badius temminckii: Starin, 2001; Japanese macaques: Takahata et al., 2002; and see for review: Moore, 1993; van Noordwijk and van Schaik, 2004), 88 but less is known concerning patterns of inbreeding avoidance between paternal relatives (but see Alberts, 1999). In this study, we showed that the probability of paternity by a dominant male decreased when he was related to the dam at R = .5 (the highest possible relatedness coefficient in our study). Smith (1995) showed in rhesus macaques that the intensity of inbreeding avoidance was directly correlated with the closeness of kinship, as in the mandrills studied here. ”
Marie Charpentier, Patricia Peignot, Martine Hossaert-McKey, Olivier Gimenez, Joanna M. Setchell, and E. Jean Wickings., 2005. “Constraints on control: factors influencing reproductive success in male mandrills (Mandrillus sphinx).” Behavioral Ecology 16:614–623]
More reference examples on incest avoidance in multiple species:
Hoier, S., 2003. “Father absence and the age of menarch, A test of four evolutionary models,” Human Nature, Vol. 14, No. 3, pp. 209–233, Walter de Gruyter, Inc., New York.
Cockburn A, Osmond HL, Mulder RA, Green DJ, Douvle MC, 2003. Divorce, dispersal and incest avoidance in the cooperatively breeding superb fairy-wren Malurus cyaneus. J Anim Ecol 185 72:189–202;
Griffin AS, Pemberton JM, Brotherton PNM, McIlrath G, Gaynor D, Kansky R, O'Riain J, Clutton-Brock TH, 2003. A genetic analysis of breeding success in the cooperative meerkat (Suricata suricatta). Behav Ecol 14:472–480;
Mateo JM, 2003. Kin recognition in ground squirrels and other rodents. J Mammal 84:1163–1181;
Pusey A, Wolf M, 1996. Inbreeding avoidance in animals. Trends Ecol Evol 11:201–206;
Stow AJ, Sunnucks P, 2004. Inbreeding avoidance in Cunningham's skinks (Egernia cunninghami) in natural and fragmented habitat. Mol Ecol 13:443–447;
Yu XD, Sun RY, Fang JM, 2004. Effect of kinship on social behaviors in Brandt's voles (Microtus brandti). J Ethol 22:17–22.
Did fossil fuel cause capitalism or did capitalism cause the creation of the technology to use fossil fuel for industrial processes? Did population start to grow in Britain before or after industrial capitalism? Why did the industrial revolution begin in Britain? Were there any precedents? Beginning before Roman Britain, this work of evolutionary sociology also looks at how Doggerland, sea-level changes accompanying ice-ages and global warming, forestation changes, malaria and plagues may have affected population movement, along with kinship rules, inheritance laws, and access to distant and denser communities through new modes of transport. Then, departing from Roman Britain, the book examines changes to the political system, fuels, technology and demography during the Reformation, the Restoration, the Dutch capitalist revolution, and the Trade Wars, to the eve of the French Revolution, which is the subject of the next volume. Hint: The cover on this book is like a treasure map and contains the major elements of the final theory. Order Demography Territory Law2: Land-tenure and the Origins of Democracy in Britain.
Demography, Territory and Law (Volume 1: The Rules of Animal and Human Populations) identified a bio-social system that keeps populations in steady-state with their environments. In this stand-alone second volume, the author tests that theory on Britain, where the world's first remarkably fast and sustained population increase began. Did this growth coincide with disruption of clan and tribal organization and relationship to place? Other possible causes investigated include capitalism itself, as well as fossil fuel.
Did fossil fuel cause capitalism or did capitalism cause the creation of the technology to use fossil fuel for industrial processes? Did population start to grow in Britain before or after industrial capitalism?
The author finds that Britain's unusual population growth was built into the British land-tenure system, which caused more fertility opportunities and diverged from that of the Romans or their successors on the European continent. The author confirms the long held suspicion that this inheritance system had something to do with the development of capitalism in Britain rather than elsewhere, and this book develops a completely new theory of capitalism.
Beginning before Roman Britain, this work of evolutionary sociology also looks at how Doggerland, sea-level changes accompanying ice-ages and global warming, forestation changes, malaria and plagues may have affected population movement, along with kinship rules, inheritance laws, and access to distant and denser communities through new modes of transport.
The book finds that the industrial revolution was not inevitable, but more likely in Britain than elsewhere because of the confluence of land-less labour, proximity of coal and iron, and deforestation after the injection of gold and silver from the New World. As it produces more private property and capitalism, the peculiar British system increases wealth disparities and reduces democracy. In France, however, population increase and industrial capitalism did not develop spontaneously, but a democratic revolution did.
Demography, Territory and Law2: Land-tenure and the Origins of Capitalism in Britain
Fascinating and original scientific and social investigation of the origins of capitalism in Britain, using a new evolutionary sociology theory and political systems comparison (including France and Holland), with scholarly reviews of alternative theories. Explores significance of Britain's odd land-tenure and inheritance system and asks where it came from, finding answers to questions preoccupying legal and economic theoreticians since the 13th century, with a demonstration of inheritance law in Hamlet. A specialist in geopolitics and energy resources, the author weighs up the roles of different fuels and technology and the availability of labour in the British industrial revolution. Many factors impinging on Britain's unusual population growth are reviewed, including diseases, transport and fertility opportunities. Alongside economic history this complex but sparkling work chronicles changes to the environment, from climate and sea-level changes to forest cover.
There is now a long record of interventions in foreign countries that have been followed by floods of refugees. For Ethiopia modern development began at the end of the 19th century. In a recently recirculated 2008 article, Irish journalist, Kevin Myers wrote despairingly of Ethiopia's overpopulation and the hopelessness of giving more aid. "By 2050, the population of Ethiopia will be 177 million; the equivalent of France, Germany and Benelux today, but located on the parched and increasingly Protein-free wastelands of the Great Rift Valley." Myers grimly anticipates a massive overflow of emigrants with poorly nourished brains to Ireland, to America, to Australia.[1] So, why is Ethiopia in such a mess? Was it always thus?
I think that Ethiopia's problems go back to the end of the 19th century and were caused by industrialisation and furthered by 'development aid' in the cause of more industrialisation.
How long has Ethiopia been overpopulated for?
In the late 19th century Ethiopia's population was only 4 or 5 million, when Mennilik II began 'modernisation'.
After Mennilik's modernisation programs, which included massive land 'reforms' (enclosures), Ethiopia's population climbed to 10 million. In 1950, as development continued, the population climbed to 18.3 million. In 2010 it is 83 million and climbing rapidly still.
What changed?
Ethiopia is one of the oldest sites of human civilisation and international trade. It had a variety of populations within, mostly rural, including hill tribes who farmed using ploughs. The various tribal populations mostly kept to their traditional areas and were sustainable within them, although for a long time there have been conflicts between adjacent states and an ancient slave trade along certain routes.
Ethiopia after 'development':
In the late 19th century there were massive land 'reforms':
The land 'reforms' coincided with dispossession and 'integration' of clans and tribes with Mennelik II who was influenced by Russian imperialists. Thrown off their land - particularly those on the plains, the people drifted to the cities, which were growing as agriculture was industrialised and roads and electricity were installed.
This process destroyed local self-government and sustainable local economies where there was probably still men's and women's land, and marriage opportunities would have been kept low through kinship rules that limited approved spouses, [2], with late menarche the rule, and low fertility through local traditional birth control practices, including contraception, spousal non-cohabitation, abstinence, delay of coitus [3] and co-operative breeding [4] (where people live in an extended family around a principal fertile couple, taking the roles of servants, aunts, aunties).
As happened in the Netherlands and in Britain,[5] local populations were disorganised by classes that got control of the food production and government and, lacking the land that had once brought them independence, they were obliged to move to new settlements, with only their labour to sell. This is the kind of system where women and children become slaves to their husbands, fathers, uncles and brothers.
Irrigation was put in place and former grazing ranges were intensively cultivated, which required dispossessing the peoples that had led sustainable lives there for millenia, without overpopulation. Malaria became a huge problem, indicative of land-disturbance and new population movements.[6]
In the 1970s there were more huge land reforms.
In the 1980s there were mass government resettlements.
The 'reform' has not stopped. It has accelerated.[7]
It is not difficult to find many accounts of how foreign aid assisted in all these 'modernisation' projects that led to the dispossession of the peoples of Ethiopia and the surrounding regions and to the current overpopulation problem and enslavement of women, for which more 'development' continues to be recommended, under the malignant pretext that overpopulation was Ethiopia's eternal burden.
What is in it for the countries driving more 'development' in Ethiopia, Somalia, Eritrea, Egypt etc? One of the major reasons for foreign powers to maintain these countries in disorder is so that the foreign powers can continue to dominate the surrounding region with military forces, supposedly to safeguard international investments. The pretence is that these African countries and all those around the Red Sea and to the North of Syria need international aid and management, however it is clear to anyone aware of the petroleum in this region that Ethiopia could be a source of primary wealth, as it was for other reasons in Ancient Egyptian, Greek and Roman times. [8]
Development, War and Refugees
Australians rightly complain that their country is overpopulated.[9] They also rightly complain that refugee and asylum-seeker numbers are adding to this problem. But they fail to link these problems to the wars the Australian government backs in the regions where the asylum seekers come from. Australia, like the United States in the film, Mother of 7 Billion,[10] adds over-consumption to its overpopulation problem. To maintain our commodity production and our importation of new residents to keep our land-sales up, we rely on huge amounts of petroleum and one of the most important sources of petroleum is the region bordered by the Land of Punt.
If Somalia, Eritrea, Ethiopia and Sudan had maintained their efficient local economies, there would be no excuse for foreign troops and corporations to be in the region, but Somalia, Ethiopia, Eritrea and Sudan would probably all be wealthy through association with the oil producing countries.
Instead the oil producing countries and the regions buffering them are in constant state of flux, overpopulation, poverty, and war and the foreign powers actually rule the region. We hear of Somalian pirates, trying to get control of the oil, but we call our own pirates 'peace keepers'.
NOTES
[1] Kevin Myers, "Somalia is not a humanitarian disaster," The Irish Independent, 2008
[2] Fertility opportunities would have been kept low through kinship rules requiring endogamous marriages that reflected the local environment with up to 8 degrees of incest avoidance and other variations that greatly decreased mating opportunities. See Sheila Newman, Development, Territory, Law: The Rules of Animal and Human Populations, Countershock Press, 2013.
Details of the wide variety of these kinship impediments to marriage and fertility in Ethiopia can be found in Siegbert Uhlig, (Ed.), Encyclopaedia Aethiopica: He-N, Otto Harrassowitz GmbH & Co. KG, Germany, 2007, pp. 55, 405-406, cited by hbdchick at https://hbdchick.wordpress.com/tag/exogamy/. Although the afore-cited Encyclopaedia states that being unmarried is not an option in Ethiopia, this is contradicted by the presence of extended families. "The extended family arrangement is frequently found among Ethiopian families since children, male or female, who have married continue to live with their family. In some cases, unmarried aunts, uncles, cousins and even close family friends continue to dwell with the nuclear family. " http://www.africa-expert.com/ethiopia/family-and-housing/ftp://ftp.cgiar.org/ifpri/ElizabethB/Ethiopia%20papers/diFalco_Bulte_2013.pdf,
Mhairi A. Gibson and Eshetu Gurmu, "Land inheritance establishes sibling competition for marriage and reproduction in rural Ethiopia"http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3038728/#!po=21.4286: "We use rich anthropological, demographic, and socioeconomic data from five traditional rural villages in Ethiopia (Table S1), where wealth remains positively correlated with reproductive success (characteristic of preindustrial societies)."
Another likely indication of earlier prevalence of local birth control methods would be family-size trends in older women who may have had more links to traditional villages. The results varied according to tribal origins as well as religion. A survey in 1986 showed that childlessness varied from 20% to 3% in the earliest cohorts (women aged 59 or older) compared to 12% to 2% in the most recent cohorts (women aged 30-39). Although much of this was attributed to successful treatment of sexually transmitted diseases the article concludes that there is more to it than that.- See more at: http://www.popline.org/node/372469#sthash.OatvN3Yl.dpuf
[3] John Knowles (Lead author), Report, "History of Birth Control Methods," Katharine Dexter McCormick Library and the Education Division of Planned Parenthood Federation of America, 2012, http://www.plannedparenthood.org/files/2613/9611/6275/History_of_BC_Methods.pdf
[4] Cooperative breeding is similar to the extended families described (which existed in most pre-WW2 societies) in note [2] Traditional Ethiopian societies were comparable to traditional European ones in many ways, so I don't mean for the following references to imply that they were all hunter-gatherers, However, for some modern evolutionary research of the phenomenon in terms of inclusive benefits etc in human societies, see, for instance, Smaldino, Paul E., Newson, Lesley, Schank, Jeffrey C., Richerson, Peter J., "Simulating the Evolution of the Human Family: Cooperative Breeding Increases in Harsh Environments," http://www.plosone.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0080753 and Hill, Kim, and Hurtado, A. Magdalena, "Cooperative breeding in South American hunter–gatherers, " School of Human Evolution and Social Change, Arizona State University, PO Box 872402, Tempe, AZ 85287-2402, USA, http://rspb.royalsocietypublishing.org/content/early/2009/08/18/rspb.2009.1061.full.pdf+html
[5a] Sheila Newman, Demography, Territory, Law 2: Land-tenure and the Origins of Capitalism in Britain, Countershock Press, 2014.
[6] David N. Weil, "The Impact of Malaria on African Development over the Longue Durée,"
Brown University and NBERhttp://www.indiana.edu/~econdept/workshops/Fall_2010_Papers/Malaria_and_Early_African_Development.pdf: "The current geographic distribution of malaria impact may not accurately represent the historical burden of the disease. Packard (2007) gives numerous examples of how human activity has changed the intensity of malaria. Clearing forests and introducing irrigation often produces the marshy environment needed for breeding of Anopheles mosquitoes. When large tracts of land are consolidated and controlled by a small number of landowners, people living on these lands are often deprived of the tools for properly tending to it and for avoiding malaria infection. This problem is exacerbated when seasonal labor is used, as migrants may come from non-malarious zones and lack immunity, then return to these zones carrying malaria with them. As for Africa, he notes that “while climate and the presence of highly efficient vectors contributed to the persistence of malaria, conditions of production played an equal if not greater role." Webb (2008) similarly describes how the introduction of agriculture led to malaria endemicity in lowlands throughout the Eurasian landmass. Population density is also important to the nature of malaria burden. Packard argues that in the African contexts, low population densities associated with mobile hunter-gatherer populations would have been unable to support the P. Falciparum infection because of the speed with which the disease exits or kills the human host. The emergence of Anopheles gambiae is also believed to closely associated with the development of agriculture. For all these reasons, the current distribution of malaria in Africa today is potentially a poor proxy for the historical distribution." See also malaria references in Sheila Newman, Demography, Territory, Law 2: Land Tenure and the Origins of Capitalism in Britain, Countershock Press, 2014.
[8] "This is pure common-sense commercial logic,” ... “There are numerous discoveries along the East African Rift in other nations and also it ends basically in Yemen, which is a known oil-bearing territory." "http://www.bloomberg.com/news/2014-07-21/gazprombank-s-gpb-global-signs-oil-exploration-deal-in-ethiopia.html; Ogaden Basin in Ethiopia: http://en.wikipedia.org/wiki/Ogaden_Basin; "Africa's next Frontier - Ethiopia?" http://oilprice.com/Energy/Crude-Oil/Africas-Next-Oil-Frontier-Ethiopia.html]
[9] And some of us can see that the same thing is happening here, with 'development' as has happened to Ethiopia.
[10] I spoke at a screening of Mother of 7 billion in 2013 and was uncomfortable at some of the assumptions made in the film. Most difficult for me was the assumption that Ethiopians had always had huge families. There was seemingly no inkling that Ethiopia had ever been anything except poor and overpopulated - of the sudden leap from 4 million to 83 million. When one considers Ethiopia's remarkably long and impressive history, this is quite an oversight. It was because of this film that I started writing an article on Ethiopia's long and different past, but it has taken me until today to get this short version up.
Outside the property development and population growth lobby, very few people who are worried about population growth and high immigration appreciate the effect of endogamy (marrying within your people) and exogamy (marrying outside your people) on population size and fertility. They also don’t recognize its effect on the private amassing of wealthy estates and political power. Anyone who wants to understand modern day problems with overpopulation, poverty, and loss of democracy would do well to study this article. This article is intended to stimulate debate about democracy, wealth distribution, and overpopulation. The author invites critical comments and argument.
Article based on S.M. Newman Demography, Territory and Law: Land-tenure and the origins of capitalism in Britain, Countershock Press, 2014. and S.M. Newman Demography, Territory and Law: The Rules of Animal and Human Populations, Countershock Press, 2013. How to read the diagrams:White squares in the diagrams below indicate permitted marriages and black squares indicate forbidden marriages. White squares become black squares when someone is already married, although polygamy varies this factor. The symetrical rules for marriage to "in-laws" are indicated by mirror images, creating an overall pyramid form in the diagram of an extended family or clan.
"Endogamy" refers to marrying within one’s clan, tribe or similar social unit. "Exogamy" refers to marrying outside those units. The most extreme kinds of endogamy tend to be practiced by ancient royal clans, such as the Egyptians and the Incas, where there were sibling, father-daughter and grandfather-granddaughter marriages. Less extreme, but more common, are first and other cousin marriages, frequently practiced by nobility and other established clans and tribes. The wealthy, whether they are noble or not, tend to marry other wealthy people for similar reasons.
High Endogamy, Low Exogamy, Low Fertility
If you look at the white squares, you will see that the pharaohs of Ancient Egypt could marry their children and their grandchildren and other close relatives. The rigidity of this practice varied from pharaoh to pharaoh, and lesser relatives might also be married, however marriage within close blood relatives was encouraged.
The purpose of highly endogamous marriages is to preserve land and power within a small group of people (known as a caste). To this day, dynasties can only preserve themselves by intermarrying. Although sibling marriage and parent or grandparent marriage is widely prohibited, first cousin marriage practiced over several generations can bring about similarly close genetic inheritance.
Although this system promotes fertility, it only does so within a very limited pool of candidates. This means that dynasties are powerful but small populations, able to concentrate, conserve and control their material assets through numerous social, legal and genetic bonds.
Outside the easily recognized institutions of tribe and nobility, people in countries where tradition holds them close to the land and preserves their extended families, still tend to live near and to marry within their own class, region and culture. This is the case with most continental European countries. It has a moderating effect on fertility opportunities[1] and a strengthening effect on local self-government and democratic organization.[2]
Low Endogamy, High Exogamy, Low Fertility
There are very few white squares, so very few permitted marriages. With incest avoidance to the 8th degree fertility opportunities within a clan are very low. This is the opposite system to the Ancient Egyptian one.
In cultures, such as those of desert indigenous populations and South Korea, fertility is kept low by restricting marriage opportunities within the family and clan and relying on external opportunity where external opportunity is limited – for instance by distance. If you are a very small clan, with only your feet for transport and your activities take place many kilometers from the next clan’s location, your opportunities to meet suitable partners will be limited. Infertile environments - typically with low rainfall - make for low density populations and big spaces between clans. The difficulty of finding a mate in such circumstances is well shown in the film Ten Canoes. (Although admittedly there were canoes, their use in the film was local rather than inter-clan.)
High Endogamy, High Exogamy, High Fertility
High exogamy is well represented by the biblical laws of Leviticus 18, very influential on Western societies. See the diagram below.
Lots of white squares here mean that you can marry a lot of people in your clan. Brothers are encouraged to marry their deceased brother's wife and niece marriage is legal. The rules differ according to whether you are male or female. This was the system that accompanied the exhortation to "Go forth and multiply."
Western societies tend to follow the Leviticus pattern, although you do get legal restrictions on cousin marriage in some places (such as Illinois, in the United States) where first cousins are not allowed to marry until they are over 55 years old) and there are age restrictions and social restrictions on marriage between uncles/aunts and nieces and nephews.
Multiculturalism as high fertility exogamy and its effect on self-government and citizen capacity to organize
Extreme exogamy applies in the English speaking ‘settler states’ of the United States, Australia and Canada. The populations in these ‘settler states’ are in continual motion due to constant reorganization of suburbs and infrastructure to accommodate high rates of immigration. This people movement occurs at international, national, regional and inter-suburban and intercity level, rather reminiscent of the increased movement of molecules in a heated substance.
Because families and clans tend to be split and disorganized in these societies, the level of endogamy is reduced, despite lack of legal restrictions. Exogamy is strongly encouraged by policies of ‘multiculturalism’. People move far away from their parents and divorce, remarriage and serial families are frequent impoverishing factors. In continental Europe there just isn’t the same amount of structural turmoil. Although the first and second world wars in those areas did cause significant disturbance, the arrangement of clans and their geographic position in villages and towns persisted.
The most important thing to understand about endogamy and exogamy, however, is their role in promoting or limiting population growth. The diagrams in this article should help the reader to see what is meant by this.
Transport factors in creating the Post War Baby Boom
Another important factor already alluded to that affects these patterns in most cases is the introduction of new transport because it permits individuals to travel greater distances. Horses, camels and elephants will take people a lot further and afford them significantly more fertility opportunities than travel on foot will. Trains, cars, boats and planes multiply opportunity exponentially. Trains are associated with massive population growth, but they impose a geometrically restricted pattern. Those restrictions disappeared with the advent of cheap oil and the automobile. Without these there would not have been a post war baby boom without precedent in size.
More variations
In general high endogamy plus high incest prohibition means low fertility. It is difficult to find people who are not married already and who are not forbidden to you in marriage but who are also members of your tribe. A person in this situation might have to go quite a long way in search of a partner and it is likely that a fairly high proportion of the clans-people would die without marrying or having children.
High endogamy but low incest prohibition, where cousin marriage is frequent means high fertility. In these kinds of situations it is considered important to lock all the land up in the tribe but to have a large tribe with many workers and potential soldiers. Nonetheless there are strict boundaries. Marriage outside the tribe is rare, although usually some immigrants will be accepted into the tribe. Living examples of such tribes are the Karen, the Hutterites and some orthodox Jewish peoples.
High exogamy and high incest prohibition will tend to disperse a people so that they ultimately become unidentifiable as clan or tribe, so you won’t find many intact tribes like this. It is a major factor in the dispersal and disintegration of many previously discrete peoples after they become affected by colonization and lose their contact with their land. Examples include Australian aborigines and possibly the Dutch of the 16th and 17th century during the minor industrial revolution that occurred in the Netherlands and which entailed major population drift from country to city. The capturing of African slaves and their transport to the Americas and Pacific Islands like Haiti is another example where the transported survivors of peoples who probably had low fertility in their original tribes encountered significantly increased fertility opportunities.
Low incest prohibition and low endogamy mean that where a clan is not isolated, it has more fertility opportunities than one with stricter rules. Such patterns characterize the settler states of Canada, Australia, the United States and Britain. Usually even first cousin marriage is permitted, but families and clans are so dispersed and fragmented that marriage to members of unidentified and equally dispersed descendants of clans are common. With the very high immigration in these countries, this potentially results in huge population growth. These are synthetically structured societies. Such countries lack the capacity to organize from the bottom up that is possible in countries where several generations are embodied in clans and historically settled and networked in a particular locality within a larger polity. An example of this strong capacity to organize based on relatively natural distribution would be France or Japan. Some examples of this capacity to organize are the French Revolution, which was able to persist over several generations until a lasting republic was formed, and the German and Japanese manufacturing sectors.
There is good reason to think that variations in endogamy and exogamy are instinctive social responses to environmental fertility signals because these rules also occur in most other animals and plants, as The Rules of Animal and Human Populations explains in chapters 3 and 4 which are also published by themselves as The Urge to Disperse. In a globalized society these signals are diffuse, remote and confusing. Media and government interpretation of signals can influence false perceptions of real environments.
NOTES
Polygamy helps to make such populations larger. An exceptional case was King Abdul Aziz, who began the current Saudi kingdom in 1932 and had 44 legitimate sons by 17 wives. The Saudi royal family had more than 4000 princes and 30,000 noble relatives in 2002 and is considered the largest royal dynasty. Without the commercial industrialization of petroleum the kingdom and dynasty could never have been so powerful. Without this kind of intermarriage the Saudi clans would not have been able to maintain control over Saudi assets. Corporations and international interference would have eroded their power, as they do among ‘common people’ by keeping them disorganised.
“Ibn Saud fathered dozens of sons and daughters by his many wives. He had at most only four wives at one time. He divorced and married many times. He made sure to marry into many of the noble clans and tribes within his territory, including the chiefs of the Bani Khalid, Ajman, and Shammar tribes, as well as the Al ash-Sheikh (descendants of Muhammad ibn Abd al-Wahhab). He also arranged for his sons and relatives to enter into similar marriages. He appointed his eldest surviving son, Saud as heir apparent, to be succeeded by the next eldest son, Faisal. The Saudi family became known as the "royal family," and each member, male and female, was accorded the title amir or amira ("prince" or "princess"), respectively.
Ibn Saud died in 1953, after having cemented an alliance with the United States in 1945. He is still celebrated officially as the "Founder," and only his direct descendents may take on the title of "his or her Royal Highness." The date of his recapture of Riyadh in 1902 was chosen to mark Saudi Arabia's centennial in 1999 (according to the Islamic lunar calendar).” Wikipedia, http://en.wikipedia.org/wiki/House_of_Saud (Accessed 26 February 2013.)
[1] "Fertility Opportunity" is a phrase borrowed from anthropologist Virginia Abernethy's theory of that name.
[2] Without intergenerational organization in the form of locally organized clans, the French Revolution probably would not have occurred. It had to persist over several generations.
 Theory and tradition suggest that when individuals are raised in environments where kinship is emphasized, the likelihood of sexual relationships diminishes, thereby enhancing safety for children. Conversely, when children are cared for by strangers—where these protective relationship-mechanisms are absent—there may be increased vulnerability to sexual exploitation.
Theory and tradition suggest that when individuals are raised in environments where kinship is emphasized, the likelihood of sexual relationships diminishes, thereby enhancing safety for children. Conversely, when children are cared for by strangers—where these protective relationship-mechanisms are absent—there may be increased vulnerability to sexual exploitation. Video and text of Sheila Newman's speech at the Animal Justice Party's event, "Policy basis for Kangaroo treatment in the ACT," 5 April 2016: Harvesting, damage mitigation and culling probably actually accelerate population growth in roos because the smaller ones survive and adapt by sexually maturing earlier - which speeds up fertility turnover. Since 2003 DNA studies have shown that ACT and southern NSW roos, both male and female, migrate at significant rates and for longer distances than the ACT model assumes. Migration has probably been mistaken for fertility, rendering ACT roo counts unreliable and invalid. The ACT needs to stop culling and widen its research base to consider various genetically based algorithms that naturally restrain fertility opportunities in kangaroos.
Video and text of Sheila Newman's speech at the Animal Justice Party's event, "Policy basis for Kangaroo treatment in the ACT," 5 April 2016: Harvesting, damage mitigation and culling probably actually accelerate population growth in roos because the smaller ones survive and adapt by sexually maturing earlier - which speeds up fertility turnover. Since 2003 DNA studies have shown that ACT and southern NSW roos, both male and female, migrate at significant rates and for longer distances than the ACT model assumes. Migration has probably been mistaken for fertility, rendering ACT roo counts unreliable and invalid. The ACT needs to stop culling and widen its research base to consider various genetically based algorithms that naturally restrain fertility opportunities in kangaroos.





 Did fossil fuel cause capitalism or did capitalism cause the creation of the technology to use fossil fuel for industrial processes? Did population start to grow in Britain before or after industrial capitalism? Why did the industrial revolution begin in Britain? Were there any precedents? Beginning before Roman Britain, this work of evolutionary sociology also looks at how Doggerland, sea-level changes accompanying ice-ages and global warming, forestation changes, malaria and plagues may have affected population movement, along with kinship rules, inheritance laws, and access to distant and denser communities through new modes of transport. Then, departing from Roman Britain, the book examines changes to the political system, fuels, technology and demography during the Reformation, the Restoration, the Dutch capitalist revolution, and the Trade Wars, to the eve of the French Revolution, which is the subject of the next volume. Hint: The cover on this book is like a treasure map and contains the major elements of the final theory.
Did fossil fuel cause capitalism or did capitalism cause the creation of the technology to use fossil fuel for industrial processes? Did population start to grow in Britain before or after industrial capitalism? Why did the industrial revolution begin in Britain? Were there any precedents? Beginning before Roman Britain, this work of evolutionary sociology also looks at how Doggerland, sea-level changes accompanying ice-ages and global warming, forestation changes, malaria and plagues may have affected population movement, along with kinship rules, inheritance laws, and access to distant and denser communities through new modes of transport. Then, departing from Roman Britain, the book examines changes to the political system, fuels, technology and demography during the Reformation, the Restoration, the Dutch capitalist revolution, and the Trade Wars, to the eve of the French Revolution, which is the subject of the next volume. Hint: The cover on this book is like a treasure map and contains the major elements of the final theory.  There is now a long record of interventions in foreign countries that have been followed by floods of refugees. For Ethiopia modern development began at the end of the 19th century. In a recently recirculated 2008 article, Irish journalist, Kevin Myers wrote despairingly of Ethiopia's overpopulation and the hopelessness of giving more aid. "By 2050, the population of Ethiopia will be 177 million; the equivalent of France, Germany and Benelux today, but located on the parched and increasingly Protein-free wastelands of the Great Rift Valley." Myers grimly anticipates a massive overflow of emigrants with poorly nourished brains to Ireland, to America, to Australia.[1] So, why is Ethiopia in such a mess? Was it always thus?
There is now a long record of interventions in foreign countries that have been followed by floods of refugees. For Ethiopia modern development began at the end of the 19th century. In a recently recirculated 2008 article, Irish journalist, Kevin Myers wrote despairingly of Ethiopia's overpopulation and the hopelessness of giving more aid. "By 2050, the population of Ethiopia will be 177 million; the equivalent of France, Germany and Benelux today, but located on the parched and increasingly Protein-free wastelands of the Great Rift Valley." Myers grimly anticipates a massive overflow of emigrants with poorly nourished brains to Ireland, to America, to Australia.[1] So, why is Ethiopia in such a mess? Was it always thus? I think that Ethiopia's problems go back to the end of the 19th century and were caused by industrialisation and furthered by 'development aid' in the cause of more industrialisation.
I think that Ethiopia's problems go back to the end of the 19th century and were caused by industrialisation and furthered by 'development aid' in the cause of more industrialisation. Outside the property development and population growth lobby, very few people who are worried about population growth and high immigration appreciate the effect of endogamy (marrying within your people) and exogamy (marrying outside your people) on population size and fertility. They also don’t recognize its effect on the private amassing of wealthy estates and political power. Anyone who wants to understand modern day problems with overpopulation, poverty, and loss of democracy would do well to study this article. This article is intended to stimulate debate about democracy, wealth distribution, and overpopulation. The author invites critical comments and argument.
Outside the property development and population growth lobby, very few people who are worried about population growth and high immigration appreciate the effect of endogamy (marrying within your people) and exogamy (marrying outside your people) on population size and fertility. They also don’t recognize its effect on the private amassing of wealthy estates and political power. Anyone who wants to understand modern day problems with overpopulation, poverty, and loss of democracy would do well to study this article. This article is intended to stimulate debate about democracy, wealth distribution, and overpopulation. The author invites critical comments and argument.
 If you look at the white squares, you will see that the pharaohs of Ancient Egypt could marry their children and their grandchildren and other close relatives. The rigidity of this practice varied from pharaoh to pharaoh, and lesser relatives might also be married, however marriage within close blood relatives was encouraged.
If you look at the white squares, you will see that the pharaohs of Ancient Egypt could marry their children and their grandchildren and other close relatives. The rigidity of this practice varied from pharaoh to pharaoh, and lesser relatives might also be married, however marriage within close blood relatives was encouraged. 




Recent comments